α-Lipoic Acid Inhibits Apoptosis by Suppressing the Loss of Ku Proteins in Helicobacter pylori-Infected Human Gastric Epithelial Cells.
Study Design
- Çalışma Türü
- Other
- Popülasyon
- None
- Müdahale
- α-Lipoic Acid Inhibits Apoptosis by Suppressing the Loss of Ku Proteins in Helicobacter pylori-Infected Human Gastric Epithelial Cells. None
- Karşılaştırıcı
- None
- Birincil Sonuç
- None
- Etki Yönü
- Mixed
- Yanlılık Riski
- Unclear
Abstract
Helicobacter pylori (H. pylori) is a Gram-negative bacterium that colonizes the gastric mucosa and triggers various stomach diseases. H. pylori induces reactive oxygen species (ROS) production and DNA damage. The heterodimeric Ku70/Ku80 protein plays an essential role in the repair of DNA double-strand breaks (DSB). Oxidative stress stimulate apoptosis and DNA damage that can be repaired by Ku70/80. However, excessive reactive oxygen species (ROS) can cause Ku protein degradation, resulting in DNA fragmentation and apoptosis. α-lipoic acid (α-LA), which is found in organ meats such as liver and heart, spinach, broccoli, and potatoes, quenches free radicals, chelates metal ions, and reduces intracellular DNA damage induced by oxidative stress. Here, we investigated whether H. pylori decreases Ku70/80 and induces apoptosis, and whether α-LA inhibits changes induced by H. pylori. We analyzed ROS, DNA damage markers (γ-H2AX, DNA fragmentation), levels of Ku70/80, Ku-DNA binding activity, Ku80 ubiquitination, apoptosis indices (Bcl-2, Bax, apoptosis-inducing factor (AIF), and caspase-3), and viability in a human gastric epithelial adenocarcinoma cell line (AGS). H. pylori increased ROS, DNA damage markers, Ku80 ubiquitination, and consequently induced apoptosis. It also decreased nuclear Ku70/80 levels and Ku-DNA-binding activity; increased Bax expression, caspase-3 cleavage, and truncated AIF; but decreased Bcl-2 expression. These H. pylori-induced alterations were inhibited by α-LA. The antioxidant N-acetylcysteine and proteasome inhibitor MG-132 suppressed H. pylori-induced cell death and decreased nuclear Ku70/80 levels. The results show that oxidative stress induced Ku70/80 degradation via the ubiquitin-proteasome system, leading to its nuclear loss and apoptosis in H. pylori-infected cells. In conclusion, α-LA inhibited apoptosis induced by H. pylori by reducing ROS levels and suppressing the loss of Ku70/80 proteins in AGS cells.
Kısaca
Results show that oxidative stress induced Ku70/80 degradation via the ubiquitin–proteasome system, leading to its nuclear loss and apoptosis in H. pylori-infected cells.
Full Text
nutrients
Article
α-Lipoic Acid Inhibits Apoptosis by Suppressing the Loss of Ku Proteins in Helicobacter pylori-Infected Human Gastric Epithelial Cells
Dayong Park , Joo Weon Lim and Hyeyoung Kim *
Brain Korea 21 FOUR Project, Department of Food and Nutrition, College of Human Ecology, Yonsei University, Seoul 03722, Korea
* Correspondence: [email protected]
Citation: Park, D.; Lim, J.W.; Kim, H. α-Lipoic Acid Inhibits Apoptosis by Suppressing the Loss of Ku Proteins in Helicobacter pylori-Infected Human Gastric Epithelial Cells. Nutrients 2022, 14, 3206. https://doi.org/ 10.3390/nu14153206
Academic Editor: Andrea Fabbri
Received: 15 July 2022 Accepted: 3 August 2022 Published: 5 August 2022
Copyright: © 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https:// creativecommons.org/licenses/by/ 4.0/).
Abstract: Helicobacter pylori (H. pylori) is a Gram-negative bacterium that colonizes the gastric mucosa and triggers various stomach diseases. H. pylori induces reactive oxygen species (ROS) production and DNA damage. The heterodimeric Ku70/Ku80 protein plays an essential role in the repair of DNA double-strand breaks (DSB). Oxidative stress stimulate apoptosis and DNA damage that can be repaired by Ku70/80. However, excessive reactive oxygen species (ROS) can cause Ku protein degradation, resulting in DNA fragmentation and apoptosis. α-lipoic acid (α-LA), which is found in organ meats such as liver and heart, spinach, broccoli, and potatoes, quenches free radicals, chelates metal ions, and reduces intracellular DNA damage induced by oxidative stress. Here, we investigated whether H. pylori decreases Ku70/80 and induces apoptosis, and whether α-LA inhibits changes induced by H. pylori. We analyzed ROS, DNA damage markers (γ-H2AX, DNA fragmentation), levels of Ku70/80, Ku–DNA binding activity, Ku80 ubiquitination, apoptosis indices (Bcl-2, Bax, apoptosisinducing factor (AIF), and caspase-3), and viability in a human gastric epithelial adenocarcinoma cell line (AGS). H. pylori increased ROS, DNA damage markers, Ku80 ubiquitination, and consequently induced apoptosis. It also decreased nuclear Ku70/80 levels and Ku–DNA-binding activity; increased Bax expression, caspase-3 cleavage, and truncated AIF; but decreased Bcl-2 expression. These H. pylori-induced alterations were inhibited by α-LA. The antioxidant N-acetylcysteine and proteasome inhibitor MG-132 suppressed H. pylori-induced cell death and decreased nuclear Ku70/80 levels. The results show that oxidative stress induced Ku70/80 degradation via the ubiquitin–proteasome system, leading to its nuclear loss and apoptosis in H. pylori-infected cells. In conclusion, α-LA inhibited apoptosis induced by H. pylori by reducing ROS levels and suppressing the loss of Ku70/80 proteins in AGS cells.
Keywords: α-lipoic acid; apoptosis; Helicobacter pylori; gastric epithelial cells; Ku protein
1. Introduction
Helicobacter pylori (H. pylori) is a Gram-negative, microaerobic, spiral bacterium with a global prevalence [1]. In 2018, Zamani, et al. [2] reported that 44.9% of the world population was estimated to have been infected with H. pylori based on a meta-analysis of studies conducted from 2009 to 2016; however, the prevalence rate varies according to socioeconomic and geographic factors. H. pylori plays a pivotal role in the pathogenesis of various stomach diseases, ranging from gastritis to gastric cancer, by producing free radicals, including reactive oxygen species (ROS) and inflammatory cytokines [3–5]. Increased ROS levels in the gastric mucosa, caused by persistent H. pylori colonization [6], can lead to mutations and DNA damage, which can be predisposing factors for cancer [7,8].
ROS comprise endogenous factors that can damage DNA by oxidizing bases and causing double-strand breaks (DSBs) [9,10]. Persistent damaged DNA threatens genomic stability, which subsequently increases the likelihood of mutagenesis and carcinogenesis [11]. However, numerous pathways for repairing damaged DNAs, collectively known
Nutrients 2022, 14, 3206. https://doi.org/10.3390/nu14153206 https://www.mdpi.com/journal/nutrients
as DNA damage responses (DDRs), can repair base adductions, deletions, and single-strand breaks via base excision, nucleotide excision, and other pathways [12–14].
DSBs are repaired through homologous recombination (HR) and canonical nonhomologous end joining (NHEJ) pathways [14,15]. The HR pathway uses adjacent homologous strands as templates to restore severed double strands, whereas after overhanging nucleotides are processed, the NHEJ pathway directly ligates damaged DNA ends [16]. Although both pathways can help to prevent chromosomal changes resulting from DSBs, the NHEJ pathway is more susceptible to local chromosome rearrangements than the HR pathway because of the absence of template strands [17,18].
The Ku70/80 heterodimer is essential for DSB repair via the NHEJ pathway [19,20]. After blocking damaged DNA ends, Ku forms a protein complex with the DNA-dependent protein kinase catalytic subunit (DNA-PKcs), Artemis, XRCC4, and ligase IV to mediate the consecutive NHEJ steps involved in DSB repair [21]. In addition, the Ku protein regulates cellular processes such as cell cycle arrest, telomere maintenance, and apoptosis [22]. The Ku70 protein can inhibit apoptosis by interacting with the pro-apoptotic protein Bax and blocking its translocation into the mitochondria [23–25]. However, when the intracellular DNA damage exceeds repair capacity, the expression of Ku70 and Ku80 is suppressed and apoptosis occurs [26–30]. We previously showed that oxidative stress decreases cell viability and Ku70 and Ku80 expression, indicating that such suppression plays an important role in apoptosis associated with excessive DNA damage [31]. When DSBs form, H2A histone family member X (H2AX) is phosphorylated to γ-H2AX, resulting in the formation of discrete γ-H2AX foci at DSBs [32].
α-lipoic acid (α-LA) is an organosulfur compound derived from octanoic acid. It contains two sulfur atoms connected by a disulfide bond. The carbon atom at c6 is chiral and the molecule exits as two enatiomers, (R)-(+)-LA and (S)-(-)-LA. Only (R)-(+)-LA (Rα-LA) exists in nature and is essential for aerobic metabolism [33]. Animal products such
- as red meat and organ meats are great sources of α-LA, but plant foods such as broccoli, tomatoes, spinach, and Brussels sprouts also contain it. [34]. In small amounts, α-LA can be enzymatically synthesized from octanoic acid in the mitochondria of humans [35]. α-LA plays an antioxidant role by quenching free radicals, chelating metal ions, and regenerating other antioxidants [36]. Owing to its powerful antioxidant properties, α-LA has recently been highlighted as a potential treatment for various diseases such as type-2 diabetes mellitus, Alzheimer disease, and many types of cancer [37,38]. α-LA protects human amniotic cells and DNA from hydrogen peroxide [39], and as well as the rat hippocampus against developing ethanol-induced DNA damage [40]. However, the precise mechanism through which α-LA protects cells against DNA damage remains unclear.
The present study aimed to determine whether H. pylori induces apoptosis by decreasing Ku70/80 proteins and whether α-LA inhibits H. pylori-induced DNA damage and apoptosis by reducing ROS levels and suppressing the loss of Ku70/80 in human gastric epithelial AGS cells.
We incubated AGS cells with antioxidant N-acetylcysteine (NAC) or the MG-132 proteasome inhibitor, and then stimulated them with H. pylori to determine the involvement of ROS and proteasomal degradation pathways in H. pylori-induced apoptosis.
For DNA markers and apoptosis indices, pro-apoptotic protein Bax, an anti-apoptotic protein Bcl-2, DNA fragmentation, caspase-3 cleavage, apoptosis inducing factor (AIF) activation, and H2AX focus formation were determined.
2. Materials and Methods
- 2.1. Cell line and Culture Conditions
Human gastric epithelial AGS cells (American Type Culture Collection, Rockville, MD, USA) were seeded in a complete medium comprising Roswell Park Memorial Institute (RPMI) 1640 medium (GIBCO, Grand Island, NY, USA) supplemented with 10% fetal bovine serum (FBS), 2 mM glutamine, 100 units/mL penicillin, and 100 µg/mL streptomycin
(Sigma-Aldrich Corp., St. Louis, MO, USA), and were then cultured at 37 ◦C under a humidified 5% CO2 atmosphere.
- 2.2. Bacterial Strain and Growth Conditions
- 2.3. Reagents
- 2.4. Infection of AGS Cells with H. pylori
- 2.5. Experimental Protocols
- 2.6. Preparation of Whole-Cell Extracts and Nuclear Extracts
Cells (1.0 × 106/10 mL) were harvested using trypsin/EDTA, followed by centrifugation at 1000× g for 5 min. Cell pellets were resuspended in 100 µL buffer containing 10 mM Tris pH 7.4, 15 mM NaCl, 1% NP-40, and 1 tablet/50 mL protease inhibitor complex (Complete protease inhibitor; Roche, Mannheim, Germany), then lysed using a 1 mL syringe with several rapid strokes. The lysate was placed on ice for 30 min, followed by centrifugation at 13,000× g for 15 min. Supernatants were collected as whole-cell extracts. Nuclear extracts were prepared using an NE-PER Nuclear and Cytoplasmic Extraction Kit (Thermo Fisher Scientific Inc., Waltham, MA, USA), as described by the manufacturer. Briefly, the cells suspended in a cytoplasmic extraction reagent containing protease inhibitor complex were vortex-mixed for 15 s, then centrifuged at 13,000× g for 10 min. Nuclear pellets were
resuspended in a nuclear extraction reagent, vortex mixed, and centrifuged at 13,000× g for 10 min. The supernatants were collected as nuclear extracts, the specificity of which was confirmed by identifying lamin B1 in the nuclear extracts. The protein concentrations were determined using Bradford assays (Bio-Rad Laboratories Inc., Hercules, CA, USA).
- 2.7. Measurement of Cell Viability and DNA Fragmentation
- 2.8. Measurement of Intracellular ROS Levels
- 2.9. Electrophoretic Mobility Shift Assay (EMSA)
- 2.10. Western Blotting
- 2.11. Immunofluorescence Staining
The cells (2.0 × 105/2 mL) were fixed in 4% formaldehyde and incubated with a blocking buffer containing 1% BSA and 0.1% gelatin for 1 h, followed by anti-γ-H2AX antibody for 1 h. After three washes with PBS, the cells were incubated with rhodamineconjugated goat anti-rabbit IgG antibody (Santa Cruz Biotechnology) for 1 h. The cells
were washed again, incubated with 5 µg/mL 4 ,6-diamidino-2-phenylindole (DAPI) in a blocking buffer for 30 min, and covered with Vectashield antifade medium. Images of the cells stained with rhodamine and DAPI were acquired using a laser scanning confocal microscope (Zeiss LSM 900; Carl Zeiss AG, Jena, Germany).
- 2.12. Co-Immunoprecipitation Assay
- 2.13. Statistical Analysis
All of the data were statistically analyzed using one-way analysis of variance (ANOVA) followed by Tukey’s post hoc test and are reported as means ± standard error of three independent experiments (n = 4 per group). Values with p < 0.05 were considered statistically significant.
3. Results
- 3.1. H. pylori Induces DNA Fragmentation and Apoptosis in AGS Cells
- 3.2. α-LA Inhibits H. pylori-Induced DNA Fragmentation and Apoptosis in AGS Cells
- 3.3. α-LA Inhibits H. pylori-Induced Increases in ROS and γ-H2AX in AGS Cells
H. pylori increased ROS levels at 1 h, and even more at 2 h (Figure 3A). Thus, the effects of α-LA on ROS levels were determined in cells stimulated with H. pylori for 2 h (Figure 3B). α-LA decreased the levels of ROS and γ-H2AX increased by H. pylori (Figure 3C).
- Nutrients 2022, 14, 3206 6 of 13
Figure 1. H. pylori increases the rates of cell death, DNA fragmentation, and apoptosis indices (Bcl-2, Bax, caspase-3, and AIF) in AGS cells. (A). The cells were stimulated without ( ) or with ratios of H. pylori to bacteria of 20:1 ( ), 50:1 (•), and 100:1 ( ), for up to 24 h. Viable cells were determined by trypan blue exclusion. All data are shown as means ± S.E. of three independent experiments.
- * p < 0.05 vs. unstimulated cells (None). (B) Cells were stimulated with H. pylori at indicated ratios of bacteria to cells for 24 h. DNA fragmentation, expressed as an enrichment factor, of unstimulated cells (None) was set at 1. All data are shown as means ± S.E. of three independent experiments.
- * p < 0.05 vs. unstimulated cells (None). (C) The cells were stimulated with 100:1 ratio of H. pylori to cells for indicated periods. Levels of Bcl-2, Bax, caspase 3, cleaved caspase 3, and truncated AIF in whole-cell extracts were determined by Western blotting. Actin was the loading control. AIF, apoptosis-inducing factor.
Using a confocal laser scanning microscope to visualize the intracellular γ-H2AX as the fluorescence intensity of rhodamine attached to an anti-γ-H2AX antibody, the status of γ-H2AX focus formation was determined. Cells stimulated with H. pylori formed robust γ-H2AX foci (Figure 3D, panel 2), which were decreased by α-LA (Figure 3D, panel 3). These results indicate that α-LA inhibited oxidative DNA damage via the antioxidant activity in the cells stimulated with H. pylori.
3.4. α-LA Inhibits H. pylori-Induced Decreases in Nuclear Ku70/80 in AGS Cells
Figure 4A shows that H. pylori decreased Ku proteins in the nuclear extracts. α-LA prevented a reduction in nuclear Ku70/80 induced by H. pylori in AGS cells (Figure 4A).
For determining the Ku–DNA binding activity, nuclear extracts were incubated with 32P-labeled Ku oligonucleotides, resolved by electrophoresis, and exposed to radiography film. As shown in Figure 4B, H. pylori decreased the Ku–DNA binding activity in the nuclear extracts, which was inhibited by the treatment of α-LA.
To determine the mechanism underlying the decrease in Ku, co-immunoprecipitation assays of the interactions between Ku80 and ubiquitin were performed. As shown in Figure 4C, H. pyori stimulation significantly increased the levels of ubiquitin-associated Ku80. Each ubiquitin molecule added ~8.5 kDa to the target protein Ku80, so an increased molecular weight of the protein was visible in the form of one or more higher molecular bands when stained with the Ku80 antibody. The results indicate that Ku80 is subject to poly-ubiquitination. The typical “ladder” of ubiquitinated Ku80 was higher than 150 kDa in size in the present study. H. pylori-induced ubiquitination of Ku80 was suppressed by α-LA.
- Nutrients 2022, 14, 3206 7 of 13
- Figure 2. α-LA inhibits H. pylori-induced cell death, DNA fragmentation, and apoptosis indices (Bcl-2, Bax, caspase-3, and AIF) in AGS cells. The cells were incubated with indicated concentrations of α-LA for 2 h, and stimulated for 24 h with an H. pylori ratio to cells of 100:1. (A) Viable cells were evaluated by trypan blue exclusion. (B). DNA fragmentation, expressed as an enrichment factor, of unstimulated cells (None) was set at 1. All of the data are shown as means ± S.E. of three independent experiments. * p < 0.05 vs. unstimulated cells (None). + p < 0.05 vs. control cells (stimulated with H. pylori without α-LA). (C) Western blots of Bcl-2, Bax, caspase-3, cleaved caspase-3, and AIF proteins in whole-cell extracts. Actin was the loading control. AIF, apoptosis-inducing factor.
- Figure 3. α-LA suppresses the H. pylori-induced increase in ROS levels, H2AX phosphorylation, and γ-H2AX foci formation in AGS cells. (A) Cells stimulated with an H. pylori ratio to cells of 100:1 for up to 4 h. The cells were incubated with α-LA at indicated concentrations for 2 h then stimulated with an H. pylori ratio to cells of 100:1 for 2 (B) and 24 (C,D) h. (A,B) ROS levels shown were determined using 2 ,7 -dichlorofluorescin diacetate (DCFH-DA) assays. All of the data are presented as means ± S.E. of three independent experiments. * p < 0.05 vs. none (unstimulated cells without α-LA); + p < 0.05, vs. Control (cells stimulated by H. pylori without α-LA). (C) Western blots of γ-H2AX in whole-cell extracts. Actin was the loading control. (D) Cells and nuclei were stained with anti-γ-H2AX antibody (red) and DAPI (blue), respectively.
- 3.5. Both NAC and MG-132 Inhibited Cell Death and Loss of Nuclear Ku Induced by H. pylori in AGS Cells
Figure 5A shows that NAC and MG-132 protected against cell death induced by H. pylori stimulation. Furthermore, a decrease in nuclear Ku70/80 induced by H. pylori was inhibited by NAC and MG-132 (Figure 5B). These results suggest that H. pylori-induced cell death and the loss of nuclear Ku70/80 are mediated by ROS and the ubiquitin-proteasome system in AGS cells.
- Figure 4. α-LA inhibits H. pylori-induced decreases in the nuclear Ku70/80, Ku–DNA binding activity, and ubiquitination of Ku80 in AGS cells. The cells were incubated with α-LA at the indicated concentrations for 2 h, then stimulated with an H. pylori to AGS cells ratio of 100:1 for 24 h. (A) Western blots of Ku70/80 in nuclear extracts. Actin and lamin B1 served as the loading control and nuclear marker, respectively. (B) The activity of Ku binding to DNA in the nuclear extracts was determined by an electrophoretic mobility shift assay. (C) Whole-cell extracts were immunoprecipitated (IP) with the anti-ubiquitin antibody, followed by Western blot analysis (WB) with anti-Ku80 antibody. The levels of the input proteins, Ku80 and actin, were determined by Western blot analysis. Input was used as the control for the protein expression.
- Figure 5. Both NAC and MG-132 suppress H. pylori-induced cell death and loss of nuclear Ku70/80 in AGS cells. Cells were incubated with 1 mM NAC or 0.5 µM MG-132 for 1 h, then stimulated with an H. pylori to AGS cells ratio of 100:1 for 24 h. (A) Viable cells were evaluated by trypan blue exclusion. All data are shown as means ±S.E. of three independent experiments. * p < 0.05 vs. None (unstimulated cells without NAC or MG-132). + p < 0.05 vs. Control (H. pylori-stimulated cells without NAC or MG-132). (B) Western blots of Ku70/80 in nuclear extracts with actin and lamin B1 as loading control and nuclear marker, respectively.
4. Discussion
As summarized in Figure 6, the present study shows that H. pylori increases ROS, which induces the degradation of Ku70/80 via the ubiquitin–proteasome system, resulting in a loss of Ku70/80 in H. pylori-infected cells. H. pylori increases oxidative DNA damage, determined by the DNA fragmentation and H2AX phosphorylation in AGS cells. Loss of Ku proteins decreases the DNA repair process and apoptosis, assessed by increased Bax/Bcl-2, caspase-3 cleavage, truncated AIF, and cell death, in H. pylori-infected cells. α-LA reduces ROS levels and thus inhibits Ku degradation, loss of Ku70/80, DNA fragmentation, and the apoptosis of H. pylori-infected cells. The antioxidant NAC and proteasome inhibitor MG-132 suppresses the degradation of Ku70/80 and apoptosis in H. pylori-stimulated cells. The results show that oxidative stress might induce Ku70/80 degradation via the ubiquitin–proteasome system, causing a loss of Ku70/80 and apoptosis of the cells infected with H. pylori. α-LA inhibits the ROS-mediated degradation of Ku70/8 and DNA damage, which suppresses apoptosis in H. pylori-infected cells. Therefore, the dietary intake of α-LA-rich foods or the supplementation of α-LA may prevent H. pylori-associated gastric cancer development and progression.
- Figure 6. The proposed mechanism through which α-LA inhibits the loss of Ku proteins and apoptosis in H. pylori-infected cells. H. pylori increases ROS, which induces the degradation of Ku70/80 via the ubiquitin–proteasome system, resulting in the loss of nuclear Ku70/80 in H. pylori-infected cells. H. pylori increases oxidative DNA damage, determined by DNA fragmentation and H2AX phosphorylation in AGS cells. Loss of Ku proteins decreases the DNA repair process and induces apoptosis, assessed by increased Bax/Bcl-2, caspase-3 cleavage, truncated AIF, and cell death, in H. pylori-infected cells. α-LA reduces ROS levels and inhibits Ku degradation, loss of Ku70/80, DNA fragmentation, and apoptosis of H. pylori-infected cells. The antioxidant N-acetylcysteine and proteasome inhibitor MG-132 suppresses the degradation of Ku70/80 and apoptosis in H. pylori-infected cells. Red arrows represent the effect of α-LA. Red blocks mean “inhibition”. AIF, apoptosis-inducing factor; ROS, reactive oxygen species.
H. pylori infection is correlated with the pathogenesis of gastric cancer [41]. We previously showed that ROS produced by NADPH oxidase increase the expression of interleukin-8 and mitochondrial dysfunction to propagate ROS production in infected gastric epithelial cells [42–44]. Biopsy specimens of gastric mucosa from patients infected with H. pylori revealed increased reactive oxygen metabolite levels [45]. As intracellular ROS accumulation induces DNA damage, oxidative stress can lead to apoptosis of gastric epithelial cells infected with H. pylori [46].
Extrinsic and intrinsic apoptotic stimuli upregulate the activity of pro-apoptotic Bcl-2 families such as Bax and Bak, and downregulate that of anti-apoptotic Bcl-2 families such
as Bcl-2 and Bcl-XL. Proteins released from the intermembrane space of the mitochondria lead to the activation of caspase-3 [47,48]. Caspase-3 is the executioner of apoptosis, and its activated form has proteolytic activities against various important proteins, leading to chromatin condensation, cytoskeletal destruction, and DNA fragmentation [49,50]. However, caspase-3 is not responsible for apoptotic cell death, as evidenced by the finding that breast cancer cells with a deleted caspase-3 gene still undergo apoptosis in vitro [51,52]. Caspase-3 independent apoptotic pathways are mediated by released AIF from the intermembrane space of the mitochondria, which also results in DNA fragmentation [53].
We found that caspase-3 was cleaved after 24 h of H. pylori infection. However, cell death was already evident at 12 h. Therefore, H. pylori-induced apoptosis might involve a caspase-3-indendent apoptosis pathway, such as AIF activation. After 12 h of incubation, AIF was truncated after H. pylori infection, which might have been responsible for the decreased cell viability in the present study. These results agree with the findings that the levels of truncated AIF protein increase before caspase-3 activation in H. pylori stimulated gastric epithelial cells [54,55]. Taken together, these results indicate that an increase in ROS within 2 h of H. pylori infection results in the release of truncated AIF and the activation of caspase-3 in AGS cells.
For the expression analysis of the DDR proteins, H. pylori downregulates the gene expression associated with the repair of the base and nucleotide excisions and mismatches, and the HR pathway [56]. However, whether H. pylori downregulates the NHEJ pathway has remained unclear. It is evident that oxidative stress induces apoptosis and downregulates Ku70/80 and NHEJ proteins [28–31]. Ku70 is considered to play a vital role in the maintenance of chromosomal integrity and cell survival. The positive correlation between Ku70 and cancer [57] indicates that Ku70 is an important candidate target for anti-cancer drug development. Ku70 is able to suppress apoptosis by sequestering Bax in colon cancer cells. In addition, loss of Ku80 was found in colon cancer cells [58]. Epigallocatechin-3gallate (EGCG)-induced apoptosis in lung cancer cells is mediated by decreased Ku70, and EGCG disrupts the interaction between Ku70 and Bax, which leads to an increased Bax expression in lung cancer cells [29]. These studies indicate the protective role of Ku70/80 against cancer cell apoptosis.
We previously found that ROS activates caspase-3, which induces Ku protein degradation and apoptosis in pancreatic acinar cells [30]. High levels of ROS activate caspase-3, resulting in a loss of Ku70/80 and apoptosis in gastric cancer cells [31]. These studies support the present finding showing that the loss of Ku proteins might be mediated by ROS and ubiquitin−proteasomes, leading to apoptotic cell death in H. pylori-infected cells.
As α-LA has a powerful antioxidant property, it reduces ROS and thus prevents ROSmediated DNA damage in vitro and in vivo [39,40]. In addition, α-LA reduces the levels of 8-hydroxy-2 -deoxyguanosine (8-OH-dG), a biomarker of oxidative damage, in calf thymus models of oxidative DNA damage in vitro [59]. α-LA reduces oxidative stress and DNA damage in mice with ulcerative colitis [60]. Here, we found the mechanism through which α-LA protects cells against DNA damage. α-LA inhibits ROS-mediated activation of the ubiquitin–proteasome system and the degradation of Ku70/80 in H. pylori-infected cells. Therefore, α-LA prevents H. pylori-induced loss of Ku proteins and apoptosis in gastric epithelial cells.
In conclusion, α-LA inhibits H. pylori-induced apoptosis by reducing ROS levels and preventing Ku70/80 loss and DNA damage in gastric epithelial AGS cells. Supplementation with α-LA or the consumption of α-LA-rich foods may prevent H. pylori-induced gastric cancer development by preventing the loss of Ku proteins and apoptotic cell death.
Author Contributions: Conceptualization, H.K.; funding acquisition, H.K.; supervision, H.K.; performing experiments and formal analysis, D.P.; new reagents and analytical tools, J.W.L.; writing—original draft preparation, D.P.; reviewing and editing, H.K. All authors have read and agreed to the published version of the manuscript.
Funding: This study was supported in part by the BK21 FOUR Project at Yonsei University, Korea.
Institutional Review Board Statement: Not applicable. Informed Consent Statement: Not applicable. Data Availability Statement: All data generated or analyzed during this study are included in this published article. Conflicts of Interest: The authors declare no conflict of interest.
Figures
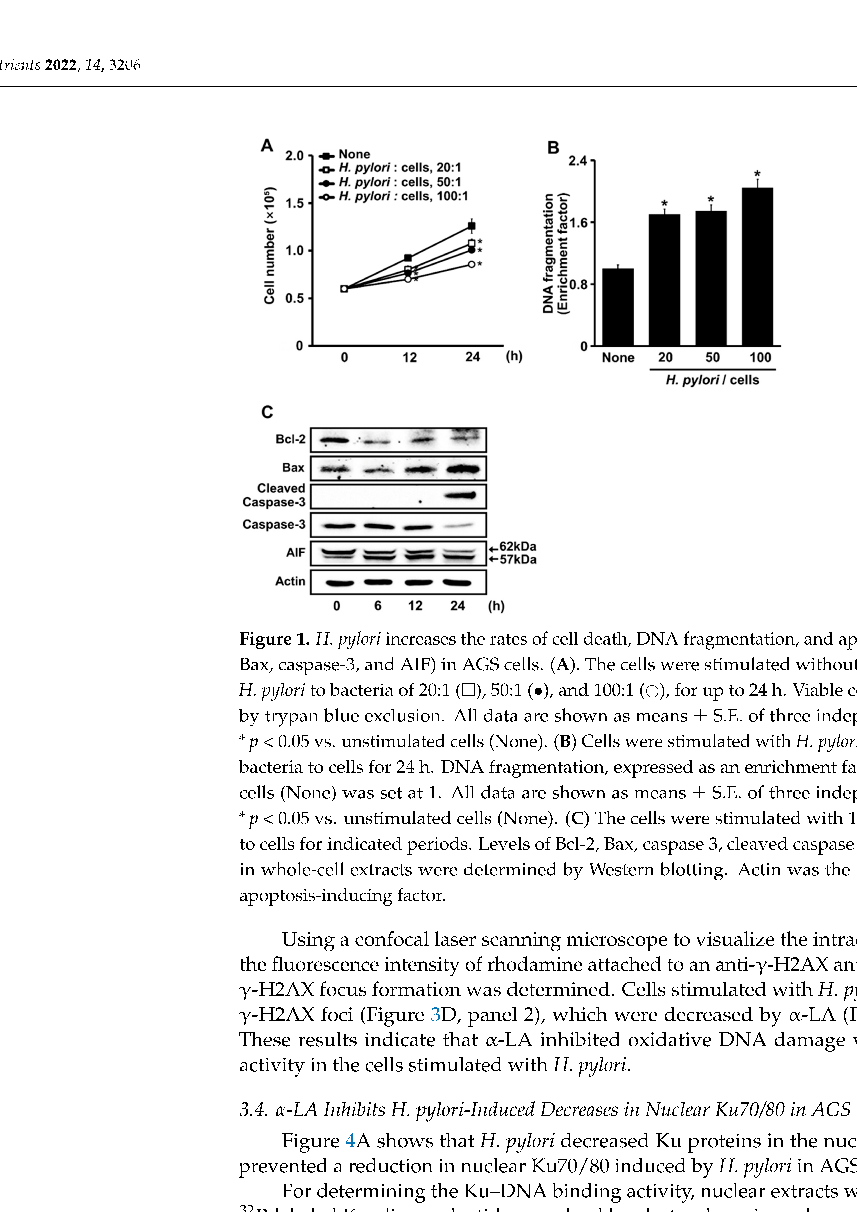
Figure 6
Immunofluorescence imaging of Ku70/Ku80 localization in gastric epithelial cells, showing nuclear protein retention in alpha-lipoic acid-treated cells despite H. pylori infection.
micrograph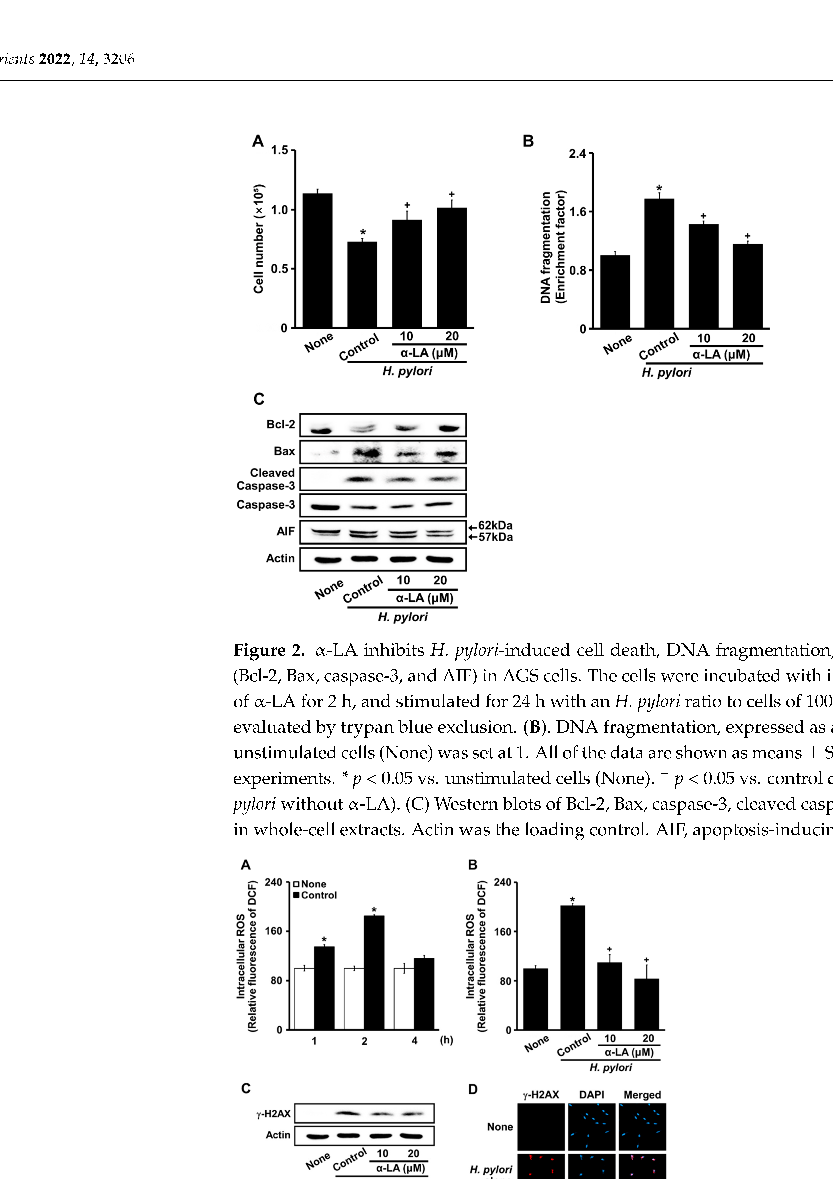
Figure 7
Quantification of DNA double-strand break repair efficiency in H. pylori-infected gastric cells, indicating that alpha-lipoic acid supports the Ku-dependent non-homologous end joining pathway.
chart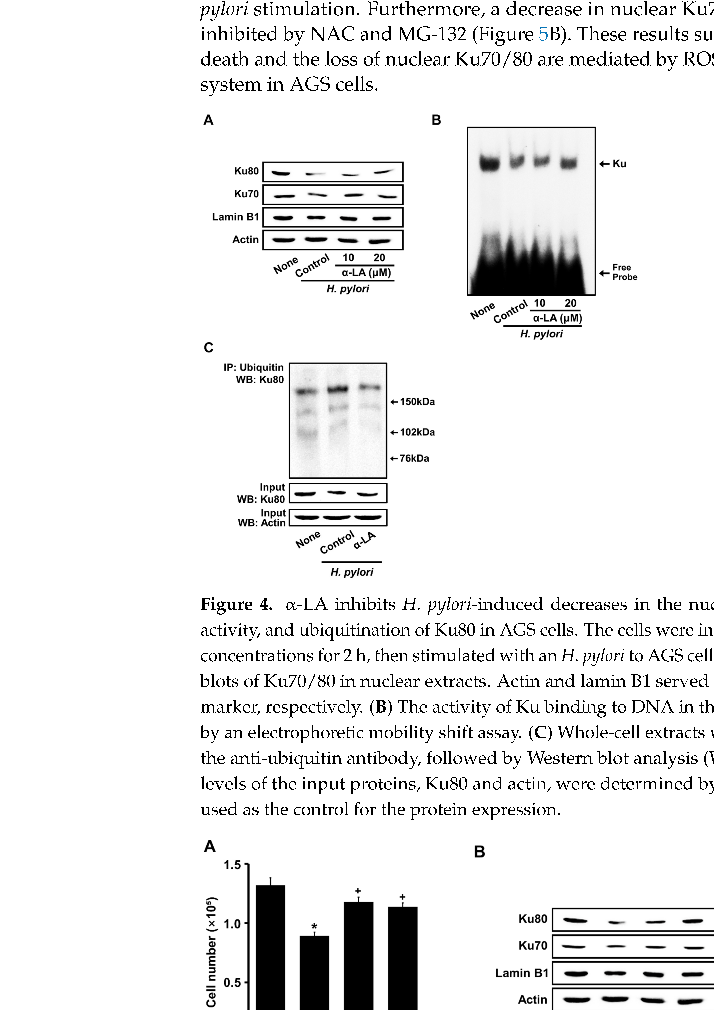
Figure 8
Cell viability analysis across different alpha-lipoic acid concentrations in H. pylori-infected gastric epithelial cells, establishing the effective dose range for cytoprotection.
chart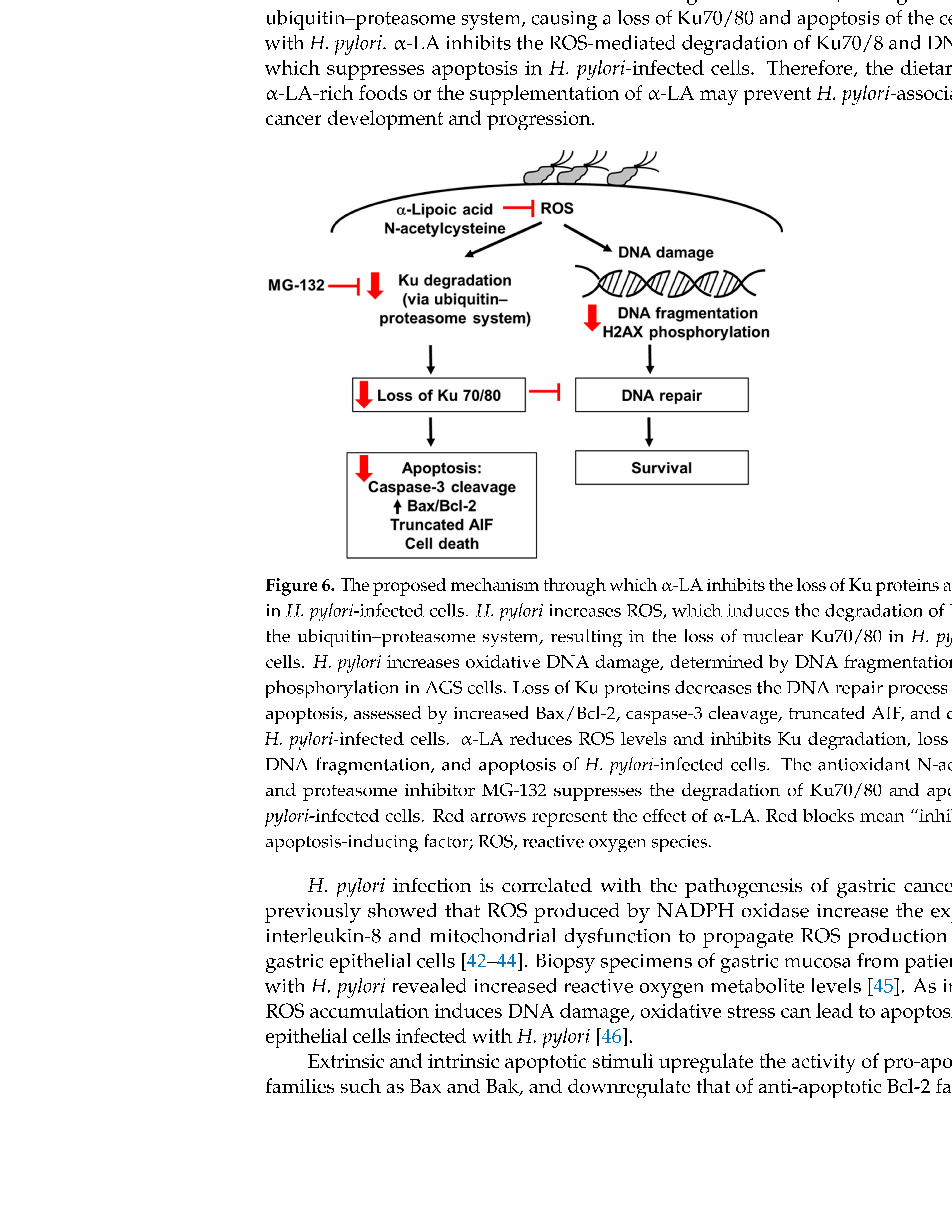
Figure 9
Signaling pathway analysis showing the molecular mechanism by which alpha-lipoic acid preserves Ku protein levels and inhibits apoptosis in H. pylori-infected gastric cells.
diagramUsed In Evidence Reviews
Similar Papers
Digestion · 2016
Reduced Abundance of Butyrate-Producing Bacteria Species in the Fecal Microbial Community in Crohn's Disease.
Gut · 1996
Evaluating the antioxidant potential of new treatments for inflammatory bowel disease using a rat model of colitis.
Journal of nanobiotechnology · 2022
Oral administration of turmeric-derived exosome-like nanovesicles with anti-inflammatory and pro-resolving bioactions for murine colitis therapy.
Frontiers in endocrinology · 2024
The emerging role of oxidative stress in inflammatory bowel disease.
The British journal of nutrition · 2012
Omega-3 fatty acids and inflammatory bowel diseases - a systematic review.
Helicobacter · 2007