The Effect of Psyllium Husk on Intestinal Microbiota in Constipated Patients and Healthy Controls.
Study Design
- Jenis Studi
- Randomized Controlled Trial
- Populasi
- healthy volunteers
- Durasi
- 1.0 weeks
- Intervensi
- The Effect of Psyllium Husk on Intestinal Microbiota in Constipated Patients and Healthy Controls. None
- Pembanding
- Placebo
- Luaran Utama
- Constipation symptoms
- Arah Efek
- Mixed
- Risiko Bias
- Low
Abstract
Psyllium is a widely used treatment for constipation. It traps water in the intestine increasing stool water, easing defaecation and altering the colonic environment. We aimed to assess the impact of psyllium on faecal microbiota, whose key role in gut physiology is being increasingly recognised. We performed two randomised, placebo-controlled, double-blinded trials comparing 7 days of psyllium with a placebo (maltodextrin) in 8 healthy volunteers and 16 constipated patients respectively. We measured the patients' gastrointestnal (GI) transit, faecal water content, short-chain fatty acid (SCFA) and the stool microbiota composition. While psyllium supplement had a small but significant effect on the microbial composition of healthy adults (increasing Veillonella and decreasing Subdoligranulum), in constipated subjects there were greater effects on the microbial composition (increased Lachnospira, Faecalibacterium, Phascolarctobacterium, Veillonella and Sutterella and decreased uncultured Coriobacteria and Christensenella) and alterations in the levels of acetate and propionate. We found several taxa to be associated with altered GI transit, SCFAs and faecal water content in these patients. Significant increases in three genera known to produce butyrate, Lachnospira, Roseburia and Faecalibacterium, correlated with increased faecal water. In summary, psyllium supplementation increased stool water and this was associated with significant changes in microbiota, most marked in constipated patients.
TL;DR
It is found that psyllium supplementation increased stool water and this was associated with significant changes in microbiota, most marked in constipated patients.
Full Text
International Journal of
Molecular Sciences
Article
The Effect of Psyllium Husk on Intestinal Microbiota in Constipated Patients and Healthy Controls
Jonna Jalanka 1,2 , Giles Major 2 , Kathryn Murray 2, Gulzar Singh 2, Adam Nowak 3, Caroline Kurtz 4, Inmaculada Silos-Santiago 5, Jeffrey M. Johnston 6, Willem M. de Vos 7 and Robin Spiller 2,*
- 1 Immunobiology Research Program and Department of Bacteriology and Immunology, Faculty of Medicine, University of Helsinki, 00014 HY Helsinki, Finland; Jonna.Jalanka@helsinki.fi
- 2 Nottingham Digestive Diseases Centre and NIHR Nottingham Biomedical Research Centre at Nottingham University Hospitals NHS Trust, the University of Nottingham, Notts NG7 2UH, UK; [email protected] (G.M.); [email protected] (K.M.); [email protected] (G.S.)
- 3 Nottingham University Hospitals NHS Trust, the University of Nottingham, Nottinghamshire NG5 1PB, UK; [email protected]
- 4 Synlogic therapeutics, Cambridge, MA 02142, USA; [email protected]
- 5 Decibel Therapeutics Inc., Boston, MA 02215, USA; [email protected]
- 6 Neurogastrx, Inc., Woburn, MA 01801, USA, formerly at Ironwood Pharmaceuticals, Cambridge, MA 02142, USA; [email protected]
- 7 Laboratory of Microbiology, Wageningen University, 6708WE Wageningen, The Netherlands; [email protected]
* Correspondence: [email protected]
Received: 17 December 2018; Accepted: 17 January 2019; Published: 20 January 2019
Abstract: Psyllium is a widely used treatment for constipation. It traps water in the intestine increasing stool water, easing defaecation and altering the colonic environment. We aimed to assess the impact of psyllium on faecal microbiota, whose key role in gut physiology is being increasingly recognised. We performed two randomised, placebo-controlled, double-blinded trials comparing 7 days of psyllium with a placebo (maltodextrin) in 8 healthy volunteers and 16 constipated patients respectively. We measured the patients’ gastrointestnal (GI) transit, faecal water content, short-chain fatty acid (SCFA) and the stool microbiota composition. While psyllium supplement had a small but significant effect on the microbial composition of healthy adults (increasing Veillonella and decreasing Subdoligranulum), in constipated subjects there were greater effects on the microbial composition (increased Lachnospira, Faecalibacterium, Phascolarctobacterium, Veillonella and Sutterella and decreased uncultured Coriobacteria and Christensenella) and alterations in the levels of acetate and propionate. We found several taxa to be associated with altered GI transit, SCFAs and faecal water content in these patients. Significant increases in three genera known to produce butyrate, Lachnospira, Roseburia and Faecalibacterium, correlated with increased faecal water. In summary, psyllium supplementation increased stool water and this was associated with significant changes in microbiota, most marked in constipated patients.
Keywords: prebiotics; microbiome; ispaghula; constipation; transit
1. Introduction
Psyllium husk, derived from the seeds of Plantago ovata, consists of highly branched and gel-forming arabinoxylan, a polymer rich in arabinose and xylose which has limited digestibility in humans. However, several members of the intestinal microbiota can utilize these oligosaccharides and
Int. J. Mol. Sci. 2019, 20, 433; doi:10.3390/ijms20020433 www.mdpi.com/journal/ijms
their constituent sugars as an energy source [1–5], and therefore, psyllium can be considered to have prebiotic potential. In general, the health-promoting effects of prebiotics include supporting the growth of bacteria beneficial to the host and increasing the production of short-chain fatty acids (SCFA) such as butyrate and propionate previously shown to be positive for colonic health [6]. Another property of psyllium is that it is capable of retaining water in the small intestine, and thereby, increasing water flow into the ascending colon. The resulting increase in the fluidity of colonic content may explain the success of psyllium husk in treating constipation [7–9]. In addition to the relief of symptoms through softening of stool and increasing stool frequency, the increase of free water alters the environmental conditions of the colon.
The effects of psyllium husk on host physiology have already been described, however, the effect that it has on the microbial ecosystem has not been well characterised. The effect of psyllium on microbial composition has been studied in an in vitro model mimicking the healthy intestine where it was shown that the microbial composition was not strongly affected. [10] However, there was an indication that the bacterial fermentation of fibres increased colonic levels of SCFAs. Moreover, a recent trial observed that psyllium supplementation protected mice from colitis by reducing the inflammatory response. This response was found to be largely microbiota dependent [11].
In this study, we aimed to determine the effect of psyllium husk supplementation on the intestinal microbiota of both healthy subjects and patients with chronic idiopathic constipation using two separate randomised, placebo-controlled intervention studies. The samples for this study come from two linked, published clinical trials [12] where the effect of psyllium husk on colonic volume and intestinal water content was tested on first healthy adult subjects and then adults with chronic constipation. Here, we built on the results reported previously by concentrating on the microbial composition alterations introduced by psyllium supplementation. We aimed to understand how altered gut environmental conditions induced by psyllium husk affected the microbiota composition. Here, we showed that in healthy controls there was a very small change in the microbiota, however, there was a larger change in the microbial composition of the constipated patients.
- 2. Results 2.1. Differences Between Healthy and Constipated Patients at Baseline
The demographics of the participants (study 1: heathy subjects, study 2: adults with chronic constipation) in the two already published clinical trials [12] are shown in Table 1. There were significant differences in age and gender distribution of the participants. Therefore, the effects of psyllium husk on the intestinal microbiota in the two studies were addressed separately and microbial composition differences between the studies were only compared at baseline.
Table 1. The patient demographics and amount of short chain fatty acids and microbial content at baseline. The standard deviations for each number show in brackets.
Demographics Healthy Constipated p-Value
Age (y) 25.75 (4.16) 41.00 (15.75) 0.02 Gender (m/f) 6/2 2/14 1.50 × 10 9 Weight (kg) 73.05 (12.49) 72.16 (15.65) 0.76 Stool water content (%) 71.13 (5.99) 65.16 (6.55) 0.07
Acetate (µM/g wet faeces) 6.90 (3.72) 21.65 (3.64) 1.5 × 10−5 Propanoate (µM/g wet faeces) 5.49 (4.24) 4.70 (3.44) 0.58 Butyrate (µM/g wet faeces) 5.88 (4.39) 5.32 (4.78) 0.78
Total bacterial load (per µL of template log10) 10.47 (0.12) 10.37 (0.28) 0.36 Methanogenic archaea (per µL of template, log10) 4.66 (2.5) 6.21 (2.05) 0.16 Microbial richness 177.75 (74.82) 201.19 (57.35) 0.35 Microbial diversity 8.26(2.71) 10.25 (6.23) 0.40
The microbial composition of healthy subjects and constipated patients differed at baseline. The health status accounted for 12% of the total microbial variation (Figure 1, p = 0.006). There were 10 taxa contributing to this difference, including 14.0-fold higher abundance of Desulfovibrio and 9.4-fold higher levels of Adlercreutzia in constipated subjects compared to the healthy controls. Moreover, the levels of Sutterella and Lachnospira were significantly decreased at baseline in constipated patients (6.7 and 4.6-fold respectively; Table 2). Moreover, there were significant differences in the amount of faecal SCFAs where the constipated subjects had 2.2-fold higher level of faecal acetate than the healthy controls (Table 1).
The microbial composition of healthy subjects and constipated patients differed at baseline. The health status accounted for 12% of the total microbial variation (Figure 1, p = 0.006). There were 10 taxa contributing to this difference, including 14.0-fold higher abundance of Desulfovibrio and 9.4-fold higher levels of Adlercreutzia in constipated subjects compared to the healthy controls. Moreover, the levels of Sutterella and Lachnospira were significantly decreased at baseline in constipated patients (6.7 and 4.6-fold respectively; Table 2). Moreover, there were significant differences in the amount of faecal SCFAs where the constipated subjects had 2.2-fold higher level of faecal acetate than the healthy controls (Table 1).
Figure 1. Principal co-ordinate analysis (PCoA) of the baseline microbial differences between the
Table 2. Relative abundance of bacterial taxa showing a statistical difference between healthy and constipated patients at baseline. Fc—fold change of taxa in constipated versus healthy subjects.
Table 2. Relative abundance of bacterial taxa showing a statistical difference between healthy and constipated patients at baseline. Fc—fold change of taxa in constipated versus healthy subjects.
Taxa Constipated Healthy Fc q-Value
Taxa Actinobacteria; Actinobacteria; Actinomycetales; Actinomycetaceae; Actinobacteria; Actinobacteria; Corynebacteriales; Nocardiaceae; Rhodococcus 0.03 % 0.00 % 6.44 Actinobacteria; Coriobacteriia; Coriobacteriales; Coriobacteriaceae; Adlercreutzia 0.10 % 0.01 % 9.44 Actinobacteria; Coriobacteriia; Coriobacteriales; Coriobacteriaceae; uncultured 0.79 % 0.17 % 4.52 Firmicutes; Clostridia; Clostridiales; Christensenellaceae; Christensenella Firmicutes; Clostridia; Clostridiales; Family XIII; 0.58 % 0.16 % 3.52 Firmicutes; Clostridia; Clostridiales; Lachnospiraceae; Lachnospira 0.13 % 0.58 % 0.22 Firmicutes; Erysipelotrichia; Erysipelotrichales; Erysipelotrichaceae; Incertae Sedis 0.46 % 0.06 % 7.15 Proteobacteria; Betaproteobacteria; Burkholderiales; Alcaligenaceae; Proteobacteria; Deltaproteobacteria; Desulfovibrionales; Desulfovibrio
Actinobacteria; Actinobacteria; Actinomycetales; Actinomycetaceae; Actinomyces
0.14% 0.02% 5.65 0.01
Actinobacteria; Actinobacteria; Corynebacteriales; Nocardiaceae; Rhodococcus
0.03% 0.00% 6.44 0.03
Actinobacteria; Coriobacteriia; Coriobacteriales; Coriobacteriaceae; Adlercreutzia
0.10% 0.01% 9.44 0.00
Actinobacteria; Coriobacteriia; Coriobacteriales; Coriobacteriaceae; uncultured
0.79% 0.17% 4.52 0.04
Firmicutes; Clostridia; Clostridiales; Christensenellaceae; Christensenella
0.13% 0.02% 6.55 0.0002
Firmicutes; Clostridia; Clostridiales; Family XIII; Incertae Sedis
0.58% 0.16% 3.52 0.01
Firmicutes; Clostridia; Clostridiales; Lachnospiraceae; Lachnospira
0.13% 0.58% 0.22 0.01
Firmicutes; Erysipelotrichia; Erysipelotrichales; Erysipelotrichaceae; Incertae Sedis
0.46% 0.06% 7.15 0.01
Proteobacteria; Betaproteobacteria; Burkholderiales; Alcaligenaceae; Sutterella
0.11 % 0.01 % 14.00
0.03% 0.23% 0.15 0.00
Proteobacteria; Deltaproteobacteria; Desulfovibrionales; Desulfovibrionaceae; Desulfovibrio
Bold: effected genus level taxon.
0.11% 0.01% 14.00 0.01
Bold: effected genus level taxon.
- 2.2. Psyllium Supplement Has a Small but Significant Effect on the Microbial Composition of Healthy Adults, but no Effect on SCFAs
The temporal stability and resistance to altered environmental conditions are key characteristics of a healthy intestinal microbiota [13]. Dietary supplementation with the carbohydrate inulin has been shown to promote expansion of Bifidobacteria in the stool in two recent studies [14,15]. Inulin has a
The temporal stability and resistance to altered environmental conditions are key characteristics
been shown to promote expansion of Bifidobacteria in the stool in two recent studies [14,15]. Inulin has a structure more readily accessible to bacteria for digestion than psyllium, which in comparative studies tends to be less fermentable than gums but more fermentable that cellulose [16]. Therefore, it was not surprising that the composition of relatively few taxa was affected by the psyllium supplementation. The only detected change was a significant increase of Veillonella (fold change (fc) = 3.7, q = 0.02) and significant decrease of Subdoligranulum (fc = −2, q = 0.01, Figure 2) with a higher dose of psyllium. There was no significant change from baseline in the levels of SCFAs measured at each study point (Supplementary Table S1).
structure more readily accessible to bacteria for digestion than psyllium, which in comparative studies tends to be less fermentable than gums but more fermentable that cellulose [16]. Therefore, it was not surprising that the composition of relatively few taxa was affected by the psyllium supplementation. The only detected change was a significant increase of Veillonella (fold change (fc) = 3.7, q = 0.02) and significant decrease of Subdoligranulum (fc = −2, q = 0.01, Figure 2) with a higher dose of psyllium. There was no significant change from baseline in the levels of SCFAs measured at each study point (Supplementary Table S1).
Figure 2. Microbial changes introduced by the psyllium supplementation to the intestinal microbiota of healthy adults.
- 2.3. Intestinal Microbiota and SCFAs of Constipated Patients Were Altered With Psyllium Supplementation
- 2.3. Intestinal Microbiota and SCFAs of Constipated Patients Were Altered With Psyllium Supplementation
In constipated patients, the effect of psyllium supplementation was greater than in healthy controls. There were seven bacterial genera with significantly altered abundance when comparing the time point of psyllium supplementation to other time points (detailed in Table 3). The psyllium husk increased the levels of Lachnospira (fc = 1.71), Faecalibacterium (fc = 2.71), Phascolarctobacterium (fc = 3.62), Veillonella (fc = 2.30) and Sutterella (fc = 2.13) when compared to baseline, whereas the levels of uncultured Coriobacteria (fc = −2.36) and Christensenella (fc = −1.83) decreased. Additionally, we detected a statistically significant difference in the amount of acetate and propionate when psyllium was consumed when compared to the baseline levels (Supplementary Table S1).
In constipated patients, the effect of psyllium supplementation was greater than in healthy controls. There were seven bacterial genera with significantly altered abundance when comparing the time point of psyllium supplementation to other time points (detailed in Table 3). The psyllium husk increased the levels of Lachnospira (fc = 1.71), Faecalibacterium (fc = 2.71), Phascolarctobacterium (fc = 3.62), Veillonella (fc = 2.30) and Sutterella (fc = 2.13) when compared to baseline, whereas the levels of uncultured Coriobacteria (fc = −2.36) and Christensenella (fc = −1.83) decreased. Additionally, we detected a statistically significant difference in the amount of acetate and propionate when psyllium was consumed when compared to the baseline levels (Supplementary Table S1).
Table 3. Bacterial taxa affected by the psyllium treatment of constipated patients. Entries represent average relative abundance. Those time points with statistically significantly different bacterial abundance from the psyllium supplementation are indicated in bold (q-values below 0.05).
Table 3. Bacterial taxa affected by the psyllium treatment of constipated patients. Entries represent average relative abundance. Those time points with statistically significantly different bacterial abundance from the psyllium supplementation are indicated in bold (q-values below 0.05).
Taxa Baseline Psyllium Wash-Out Maltodextrin
Taxa Baseline Psyllium Wash-out Maltodextrin Actinobacteria; Coriobacteriia; Coriobacteriales; Coriobacteriaceae; uncultured
Actinobacteria; Coriobacteriia; Coriobacteriales; Coriobacteriaceae; uncultured
0.80% 0.34% 0.56% 0.88%
Firmicutes; Clostridia; Clostridiales; Christensenellaceae; Christensenella
0.13% 0.07% 0.08% 0.08%
Firmicutes; Clostridia; Clostridiales; Lachnospiraceae; Lachnospira
Firmicutes; Clostridia; Clostridiales; Christensenellaceae; Christensenella 0.13 % 0.08 % 0.08 % Firmicutes; Clostridia; Clostridiales; Lachnospiraceae; Lachnospira 0.11 % 0.09 % 0.13 % Firmicutes; Clostridia; Clostridiales; Ruminococcaceae; Firmicutes; Negativicutes; Selenomonadales; Phascolarctobacterium
0.11% 0.20% 0.09% 0.13%
Firmicutes; Clostridia; Clostridiales; Ruminococcaceae; Faecalibacterium
3.48% 9.43% 8.53% 7.47%
Firmicutes; Negativicutes; Selenomonadales; Acidaminococcaceae; Phascolarctobacterium
0.54% 1.95% 0.30% 0.60%
Firmicutes; Negativicutes; Selenomonadales; Veillonellaceae; Veillonella
0.05% 0.11% 0.04% 0.06%
Proteobacteria; Betaproteobacteria; Burkholderiales; Alcaligenaceae; Sutterella
0.03% 0.07% 0.02% 0.03%
Firmicutes; Negativicutes; Selenomonadales; Veillonellaceae; Veillonella 0.05 % 0.04 % 0.06 %
Bold: Time points with statistically significantly different bacterial abundance from the psyllium supplementation are indicated in bold.
- 2.4. The Impact of Increased Transit on Intestinal Microbiota
- 2.5. Associations Between Intestinal Microbiota and Faecal Water Content and SCFAs
In line with previous reports, psyllium supplementation increased the participants’ stool water content in both studies [12]. Additionally, we showed that this same measure could be associated with changes in the intestinal microbiota in both study groups. The detailed associations can be found in Supplementary Tables S2 and S3 for constipated and healthy subjects respectively. In the constipated patient cohort, we found that several Actinobacteria taxa (genera Actinomyces, Bifidobacterium, Asaccharobacter and uncultured Coriobacter) showed significant negative associations with increased stool water content. In addition, three genera known to produce butyrate, Lachnospira, Roseburia and Faecalibacterium, were significantly increased with increased faecal water content (Figure 4). In the healthy subjects, only Anaerococcus showed a negative association to faecal water content.
Short-chain fatty acids are the end products of bacterial fermentation of dietary fibres. We found a significant association between longer transit time and lower levels of SCFAs. The amounts of acetate (cor = −0.59, p = 0.0002), butyrate (cor = −0.42, p = 0.015) and propionate (cor = −0.59, p = 0.0002) all showed a significant correlation with transit time (measured with Weighted Average Position Score (WAPS) after 48 h) in constipated patients. There were no significant correlations between SCFAs and transit times in healthy subjects.
Int. J. Mol. Sci. 2019, 20, 433 6 of 12
Figure 4. Associations between faecal water content and intestinal genera in constipated subjects at all data points.
Short-chain fatty acids are the end products of bacterial fermentation of dietary fibres. We found a significant association between longer transit time and lower levels of SCFAs. The amounts of
= 0.0002), butyrate (cor = −0.42, p = 0.015) and propionate (cor = −0.59,
all showed a significant correlation with transit time (measured with Weighted Average Position Score (WAPS) after 48 h) in constipated patients. There were no significant correlations between SCFAs and transit times in healthy subjects.
Figure 4. Associations between faecal water content and intestinal genera in constipated subjects at all data points.
Additionally, we noted that the abundance of 16 bacterial genera were associated with the amount of faecal SCFAs in the constipated patients (Supplementary Table S2). A higher amount of acetate was associated with increased levels of two genera from Lachnospiraceaea family, Roseburia (q = 0.03) and Lachnospira (q = 0.03) as well as Faecalibacterium (q = 0.0001; Figure 5). Moreover, the latter two showed positive associations between the levels of propionate (Lachnospira q = 0.002, Faecalibacterium q = 0.04). In healthy subjects, there were 10 bacterial taxa with associations to the altered levels of faecal SCFAs (Supplementary Table S3). These included the positive associations of Leuconostoc (q = 0.03) with butyrate levels and the positive association of Blautia (q = 0.07) with acetate
Short-chain fatty acids are the end products of bacterial fermentation of dietary fibres. We found a significant association between longer transit time and lower levels of SCFAs. The amounts of acetate (cor = −0.59, p = 0.0002), butyrate (cor = −0.42, p = 0.015) and propionate (cor = −0.59, p = 0.0002) all showed a significant correlation with transit time (measured with Weighted Average Position Score (WAPS) after 48 h) in constipated patients. There were no significant correlations between SCFAs and transit times in healthy subjects.
- q = 0.03) and Lachnospira (q = 0.03) as well as Faecalibacterium (q = 0.0001; Figure 5). Moreover, the
- q = 0.04). In healthy subjects, there were 10 bacterial taxa with associations to the altered levels of faecal SCFAs (Supplementary Table S3). These included the positive associations of Leuconostoc (q = 0.03) with butyrate levels and the positive association of Blautia (q = 0.07) with acetate levels.
Additionally, we noted that the abundance of 16 bacterial genera were associated with the amount of faecal SCFAs in the constipated patients (Supplementary Table S2). A higher amount of acetate was associated with increased levels of two genera from Lachnospiraceaea family, Roseburia
Figure 5. Significant associations between acetate and the abundance of microbial taxa in constipated patients.
3. Discussion
3. Discussion
In this study, we utilized two clinical trials to investigate the effects of psyllium husk ingestion on the faecal microbiota composition of both healthy controls and patients with idiopathic constipation [12]. We showed that the environmental changes (change in transit time, faecal water content and the concentration of SCFAs) introduced by the psyllium husk were substantial in both patient groups. However, the healthy subjects showed very little change in their faecal microbial composition, whereas several key members of the microbial ecosystem were altered in the constipated patients.
In this study, we utilized two clinical trials to investigate the effects of psyllium husk ingestion on the faecal microbiota composition of both healthy controls and patients with idiopathic constipation [12]. We showed that the environmental changes (change in transit time, faecal water content and the concentration of SCFAs) introduced by the psyllium husk were substantial in both patient groups. However, the healthy subjects showed very little change in their faecal microbial composition, whereas several key members of the microbial ecosystem were altered in the constipated patients.
There were detectable microbial differences between healthy and constipated patients at baseline, including significantly higher levels of Desulfovibrio ssp. in the constipated patients. Interestingly, in a recent study, a member of the Desulfovibrio genus was shown to reduce transit in an in vivo model [17]. It has also been shown that this taxon produces hydrogen sulphide, which is known to decrease inflammation in the GI tract and to inhibit motility [18,19]. The association of Desulfovibrio spp. with
constipated patients is a new finding that may be followed up by studies on the relevance of this organism to GI health and its effect on orchestrating motility.
Another genus level taxon that showed significant difference between the study groups was Lachnospira, with an over four-fold higher level in the healthy than in the constipated subjects. Additionally, we detected a significant increase of this taxon in these patients after psyllium supplementation and we found positive associations between increased faecal water content and acetate levels. It is known that several members from Lachnospira genus are capable of producing lactate and acetate. Lactate may be further metabolized into butyrate or propionate. It has been shown that low concentration of butyrate contributes to constipation by inhibiting mucin secretion [20]. Therefore, decreased levels of these SCFAs produces could contribute to the disease state of constipation.
Psyllium husk has long been used as a treatment for constipation; however, the changes it produces in the composition of the intestinal microbiota have so far not been addressed. We showed that the intestinal microbiota composition of healthy individuals was less affected by the psyllium supplementation than those of the constipated patients. This is in line with previous findings on psyllium supplementation [21] as well as the general understanding where the microbial composition of healthy subjects is regarded to be resistant to environmental changes [22]. It should be noted that the constipated patients had a larger environmental change, as reflected by the change in colonic T1, an Magnetic Resonance Imaging (MRI) time constant reflecting fluidity of the colonic chyme [12], than the healthy subjects; the degree of microbial change may be a reflection of this. We showed that in both studies there was an increase of Veillonella, an organism typically more abundant in the small intestine [23]. There were also changes in other small intestinal taxa [24], such as Sutterella (belonging to the Proteobacteria phylum), which increased in abundance after psyllium supplementation in the constipated subjects. Sutterella bacteria are slow growing organisms [25,26], and therefore, their higher abundance with faster transit could be an indication that these bacteria are washed out of the small intestine and become detectable in greater numbers in faeces. Therefore, we hypothesize that the increase in both of these bacterial taxa is secondary to the supplementation of psyllium and change in the colonic environment rather than increased colonic growth of the bacteria belonging to these taxa.
The supplementation of psyllium significantly increased the numbers of commensals such as Phascolarctobacterium [27] and Faecalibacterium [28] in the constipated patients. These same organisms have also been shown to have increased abundance after consumption of a polydextrose fibre [29]; Faecalibacterium and Roseburia have also been previously associated with faster colonic transit [30] and Faecalibacterium with loose stools [31]. In our study, the increase in Faecalibacterium was also associated with higher stool water content and the increase of both Phascolarctobacterium and Faecalibacterium with larger amounts of acetate in stools. Interestingly, Faecalibacterium converts sugars and acetate into butyrate–an important molecule for intestinal health used by the colonocytes as an energy resource. The favourable change in the acetate concentration may be explained by the increase in abundance of these genera.
Another bacterial genus affected by the psyllium supplementation in the constipated patients was Christensenella, which decreased almost two-fold in abundance after psyllium treatment. Previously, Christensenella had been associated with hard stools [31], and interestingly, we were able to show a similar trend, since in our study, this genus was negatively associated with increased faecal water content.
In conclusion, we showed that consumption of psyllium husks introduced small but significant changes in the intestinal microbiota of both healthy and constipated patients. This change was more pronounced in the constipated patients. Organisms associated with intestinal short-chain fatty acid production such as Faecalibacterium ssp. were increased, an effect that suggests a potential health benefit from psyllium supplementation. Several candidate mechanisms for such an effect were present, such as changes in gut transit and increased intestinal water content. These data support potential health benefits for such mechanisms in addition to their role in the relief of constipation symptoms.
institutional and national review boards respectively and registered on www.clinicaltrials.gov
Int. J. Mol. Sci. 2019, 20, 433 8 of 12
A detailed description of these studies has been published previously [12], thus, the methodology is here only given in brief. The study 1, in adults without digestive disorders ( subject failed to provide stool samples), was a double-blinded, placebo-controlled three-period, three-treatment crossover trial. Treatments were taken in a random order, each taken for a six-day period. Subjects took 14 g of powder three times daily, either 14 g maltodextrin, 14 g Metamucil (providing 7 g psyllium), or a 50:50 mixture 7 g maltodextrin and 7 g Metamucil (providing 3.5 g psyllium). Treatments were separated by washout periods of one week. Four faecal samples were collected, one at baseline and one after each treatment period (Figure 6).
4. Materials and Methods
- 4.1. Study Design
This study consisted of two separate, published clinical trials [12] where the effect on psyllium husk on MRI parameters of gastrointestinal physiology was tested on adults without digestive disorders (study 1) and adults with chronic constipation (study 2). The protocols were approved by institutional and national review boards respectively and registered on www.clinicaltrials.gov (NCT01805999 and NCT02144376). All participants gave written informed consent.
A detailed description of these studies has been published previously [12], thus, the methodology is here only given in brief. The study 1, in adults without digestive disorders (n = 9; one subject failed to provide stool samples), was a double-blinded, placebo-controlled three-period, three-treatment crossover trial. Treatments were taken in a random order, each taken for a six-day period. Subjects took 14 g of powder three times daily, either 14 g maltodextrin, 14 g Metamucil (providing 7 g psyllium), or a 50:50 mixture 7 g maltodextrin and 7 g Metamucil (providing 3.5 g psyllium). Treatments were separated by washout periods of one week. Four faecal samples were collected, one at baseline and one after each treatment period (Figure 6).
The study 2 included patients (n = 16) suffering from chronic constipation, which was defined by meeting the Rome III criteria for either functional constipation or constipation-predominant irritable bowel syndrome. This study was a double-blind, placebo-controlled, two-period, twotreatment study, using only maltodextrin (placebo) and the higher dose (21 g/day) of psyllium. In order to avoid any carryover effects both treatment periods were preceded with ≥10 days of usual laxative use; then, 8 days without laxatives other than rescue therapy. Rescue therapy (oral bisacodyl 5 mg) was permitted in patients who had not opened their bowels for 3 days and were experiencing distressing symptoms, but not in the 48 h before stool sample collection. Four faecal samples were collected: at baseline, after each of the treatments and after the washout period in between treatments
The study 2 included patients (n = 16) suffering from chronic constipation, which was defined by meeting the Rome III criteria for either functional constipation or constipation-predominant irritable bowel syndrome. This study was a double-blind, placebo-controlled, two-period, two-treatment study, using only maltodextrin (placebo) and the higher dose (21 g/day) of psyllium. In order to avoid any carryover effects both treatment periods were preceded with ≥10 days of usual laxative use; then, 8 days without laxatives other than rescue therapy. Rescue therapy (oral bisacodyl 5 mg) was permitted in patients who had not opened their bowels for 3 days and were experiencing distressing symptoms, but not in the 48 h before stool sample collection. Four faecal samples were collected: at baseline, after each of the treatments and after the washout period in between treatments (Figure 6).
- Study 1 – Healthy controls N = 7
- Study 2 – Constipated patients N = 16
1wk 1wk
Treatment 1 Treatment 2 Treatment 3
- 4.2. Measurements of Transit (WAPS) Subjects swallowed five identical transit markers: cylinder-shaped inert capsules containing
Figure 6. Study outline. The arrows indicate faecal sample collection points.
- 0.4 mL 15 µM gadoteric acid, a positive MRI contrast agent. Ingestion was confirmed in patients by direct observation or via a time-stamped video. The weighted average position score of the transit markers on an MRI scan 24 h after ingestion (WAPS24) was calculated using the formula (sum of the segment number X the number of markers in each segment divided by the total number of segments),
4.2. Measurements of Transit (WAPS)
Subjects swallowed five identical transit markers: cylinder-shaped inert capsules containing 0.4 mL 15 µM gadoteric acid, a positive MRI contrast agent. Ingestion was confirmed in patients by direct observation or via a time-stamped video. The weighted average position score of the transit markers on an MRI scan 24 h after ingestion (WAPS24) was calculated using the formula (sum of the segment
- as described previously [32]. A higher score denotes slower whole gut transit. WAPS48 was based on the same calculation 48 h after marker ingestion. For healthy controls, transit was measured at the end of each of the three treatment periods and for constipated patients at the end of each of the two treatments.
- 4.3. Analysis from Faecal Samples
- 4.4. Statistical Analysis
The effect of psyllium was calculated for both trials separately, since there were notable demographic differences between trial populations. Comparisons between subjects from the two trials were only undertaken at baseline to elucidate differences between the groups. The baseline differences in SCFA, stool water content, qPCR results and transit times between the patient groups were calculated either with t-test (parametric data) or Wilcox test (nonparametric data). For microbial
diversity, we used the inverse Simpson’s diversity index. Correlations between these data was analysed by using Spearman correlation.
All of the 16S rRNA gene sequencing data were analysed with in-house script from the Mare r-package [35] using R (version 3.2.3). For visualization of the overall microbial composition differences
- at baseline we performed PCoA using Bray-Curtis dissimilarities. For analysing the effect of health status on the overall microbiota composition, we used a MANOVA test. Differences in bacterial abundances and associations between bacterial taxa and SCFAs, stool water content and transit times within each study were estimated with generalized linear mixed models from all available data points. In both models, the obtained read counts were modelled as a function of time point, using subject ID
- as a random factor and the total number of reads per sample as an offset, assuming negative binomial distribution. The resultant p-values were corrected for multiple comparisons with false discovery rate (FDR), and subsequent q-values bellow 0.1 were considered to be significant [40].
- at the time of the study. Other authors have nothing relevant to declare. The funders had no role in the design of the study; in the collection, analyses or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Figures
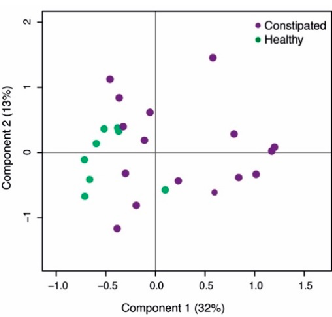
Figure 5
Principal coordinate analysis of baseline microbial communities reveals separation between constipated patients and healthy controls, indicating distinct microbiome configurations associated with chronic constipation.
chart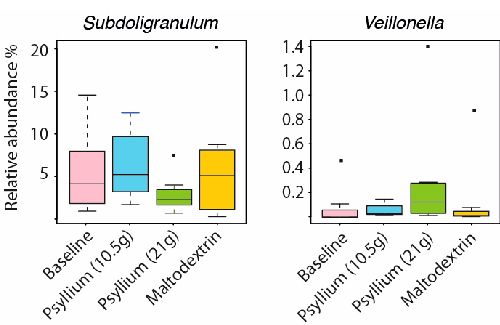
Figure 6
Psyllium supplementation alters the intestinal microbiota of healthy adults, with specific increases in Faecalibacterium and other fiber-fermenting taxa. These changes are consistent with enhanced saccharolytic fermentation.
chart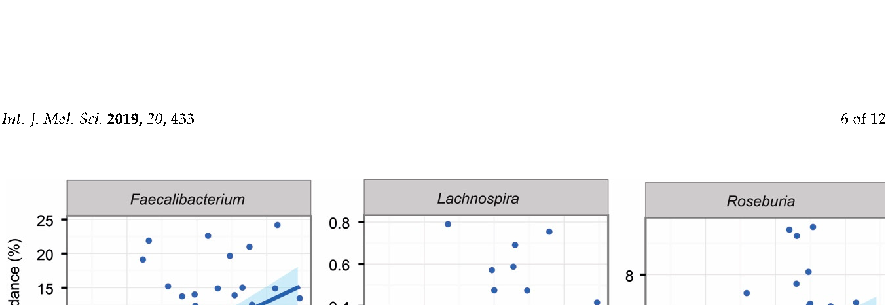
Figure 7
Microbial taxa changes in constipated patients following psyllium supplementation show distinct patterns compared to healthy volunteers. Certain Lachnospiraceae and Ruminococcaceae members increase selectively.
chart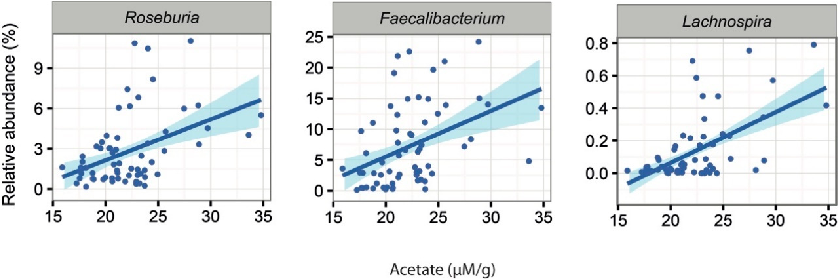
Figure 8
Significant associations between fecal acetate concentrations and specific microbial taxa are identified in constipated patients. Higher Lachnospira and Roseburia abundance correlates with increased acetate production.
chartTables
Table 1
Figure 6. Study outline. The arrows indicate faecal sample collection points.
Used In Evidence Reviews
Similar Papers
BMJ (Clinical research ed.) · 2008
Effect of fibre, antispasmodics, and peppermint oil in the treatment of irritable bowel syndrome: systematic review and meta-analysis.
Journal of the Academy of Nutrition and Dietetics · 2017
Understanding the Physics of Functional Fibers in the Gastrointestinal Tract: An Evidence-Based Approach to Resolving Enduring Misconceptions about Insoluble and Soluble Fiber.
The American journal of gastroenterology · 2013
Fiber and functional gastrointestinal disorders.
International journal of pharmaceutics · 2007
Psyllium as therapeutic and drug delivery agent.
Alimentary pharmacology & therapeutics · 2004
Systematic review: the role of different types of fibre in the treatment of irritable bowel syndrome.
BMJ (Clinical research ed.) · 2009